Trilobite Internal Anatomy
The dorsal morphology of trilobites is typically well preserved, and ventral features such as limbs and antennae are only rarely preserved. Similarly, our knowledge of the internal anatomy of trilobites is very poorly understood. X-ray images of some trilobite specimens indicate a long, central (axial) structure typically considered an alimentary canal (intestines or gut). Sometimes the gut or its contents are also preserved as an axial structure. Only in extremely well-preserved agnostid specimens has a trace of a mouth and anus been detected. The mouth is associated with the hypostome, and the anus opens toward the rear of the pygidium, as might be expected. The images below are derived from several sources, including both Treatises (1959 and 1997). Most workers rely on knowledge of living arthropod anatomy and presume that trilobites had similar (if perhaps more primitive) circulatory, nervous, and muscular systems. This leads to reconstructions as below:
..
Trilobite Internal Anatomy
This drawing ©2004 by S. M. Gon III
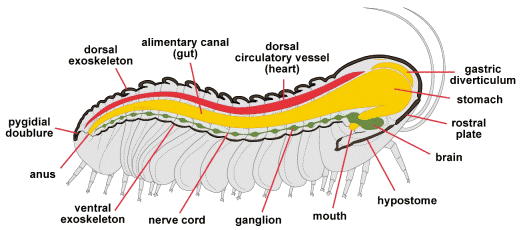
In the figure above, a longitudinal sagittal section taken through the midline of the axis of a phacopid trilobite reveals some of the major systems. The thick dorsal exoskeleton (composed of a complex structure of chitin and protein, thickened and hardened by calcite (calcium carbonate [CaCO3]) and much thinner ventral shell (uncalcified), are shown in dark brown. The dorsal circulatory vessel (heart) is shown in red. Modern arthropods have an open circulatory system in which internal organs are bathed in hemolymph (blood) which is circulated through contractions of the dorsal circulatory vessel.
The digestive system (yellow) starts at the mouth (adjacent to the hypostome), which feeds forward into a stomach which occupies much of the glabellar space. From the stomach, a long alimentary canal sends food backward to exit at the anus. Accessory digestive organs, sometimes referred to as gastric diverticulae occur both above and to each side of the stomach (see another example below).
We presume that trilobites had a ventral nervous system, either a pair of nerve cords connected at segmental ganglia (swellings), or perhaps a single cord with segmented nodes (shown above in dark green). The brain would be an enlarged frontal ganglion receiving sensory input from eyes and antennae, etc. Its precise placement is conjectural, but most modern arthropod brains are anterior to the mouth, with the paired central nerves passing to each side of the esophagus, rejoining at a subesophageal ganglion.
In the figure above, the muscular system is not shown (but see below). The reproductive system of trilobites is also very poorly understood. There is some evidence that eggs were brooded at the front of the cephalon (as they are in horseshoe crabs today), but the nature and location of copulatory organs has never been documented.
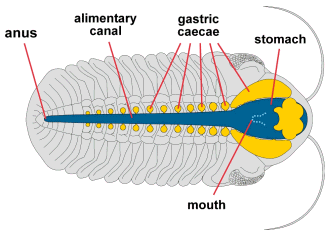
This dorsal view features the digestive system (dark blue), showing the
frontal placement of the expanded stomach, and the lateral and central
caecae (yellow), which were thought to have acted much as a liver. The
large cephalic lobe of a phacopid wasn't occupied by philosophical
considerations, but by its most recent meal! The long, median alimentary
canal (intestine) ended at a posterior anus. |
Figure after Bergstrom 1976
A complex network of subcuticular genal caecae (above) is
apparent in some trilobites (e.g., thin shelled Olenidae) and may be related to circulation or cuticular respiration. Note the patterns of outward-radiating vessels and dense palpebral anastomoses. Similar pygidial caecal networks occur in some species, indicating that they are not likely related to digestion. |
|
Trilobite cross section
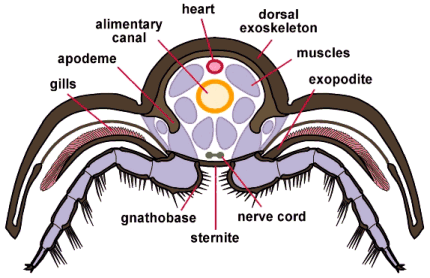
In the cross-section (left), through the thoracic region of a phacopid trilobite, we see how the organ systems were placed centrally amidst the muscular system that worked the limbs, allowed for enrollment, and changed body posture. The gills, borne on the exopodite, are shown in pink; part of the circulatory and respiratory system. Oxygen-rich hemolymph (arthropod blood) would enter the hemocoel (an open circulatory system bathing the internal organs and muscles) via the legs, and would be moved along the length of the animal via the long dorsal vessel (heart) shown here as a red oval. Below the heart is the alimentary canal (yellow) and the ventral nerve cords (olive). The muscle masses are shown in purple. The majority of organ systems occupies the axial lobe of the animal. The pleural exoskeleton provided protection for the limbs and gills, but the "guts" of a trilobite occupied the axial lobe. Some workers speculate that the excretory system of trilobites might have occupied the pleurae, exiting as ducts (associated with the Panderian organs) where the pleural doublure meets the thin ventral cuticle. Finally, the spines on the limbs, especially heavy on the coxae (base segments), were acted as teeth and jaws, breaking up food before being passed forward to the hypostome and mouth. This "jaw at the base" of the leg is called a gnathobase (translating literally as "jaw base").
|
|
As a last image, only rarely are internal features of trilobites preserved, as in this specimen showing the
digestive tract and lateral caecae of Selenocoryphe platyura, a ptychoparioid trilobite from Utah. Note
how the caecae become less prominent posteriorly, and disappear altogether by the middle of the thorax.

The Pygidium
The pygidium is a tail piece composed of a variable number of fused segments at the posterior end of a trilobite. While the cephalon of a trilobite is typically wider than the thorax, and one of the single largest major features of a trilobite body, the pygidium can range from extremely small to larger than the cephalon. There are four general categories of pygidium relative size, shown below:
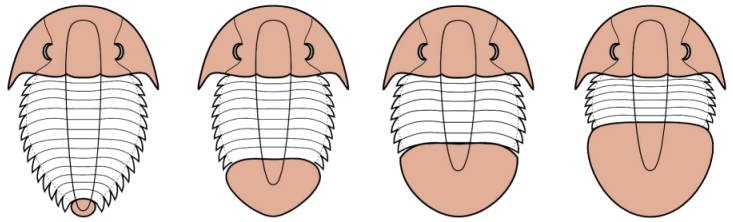 |
micropygous
pygidium much smaller
than cephalon |
subisopygous
pygidium subequal
to cephalon |
isopygous
pygidium equal
to cephalon |
macropygous
pygidium larger
than cephalon |
Above are four hypothetical examples, with cephalon and pygidium tinted
©2005 by S. M. Gon III. Created using Macromedia Freehand
The relationship between the size of the pygidium relative to the cephalon has some utility in considering trilobite classification. For example,
Redlichiida typically have
micropygous pygidia, while those of
Asaphida are typically
subisopygous to
isopygous. Thus, these terms appear in the
Trilobite Order Fact Sheets when describing the pygidium characters of the trilobite orders. Examining the subisopygous and isopygous specimens below, it is clear how the pygidium is made up of several segments, but these are fused, not articulated as thoracic segments are. Even a micropygous pygidium is typically made up of a few fused segments. The relationship between segments and articulations is discussed further in the
ontogeny and development page, where the role of the pygidium in growth and addition of segments is described. In trilobites with larger pygidia, it functions as part of a protective capsule when the trilobite
enrolls, typically matching the cephalon more or less precisely.
©2005 by S. M. Gon III. Created using Macromedia Freehand
In addition to size, the pygidium can be small and simple; or larger, but otherwise similar to the thoracic segments before it (the homonomous condition), or distinguished by modifications such as spines and lobes, or otherwise quite different in morphology from the thoracic segment pattern (the heteronomous condition). The examples below display both homonomous and heteronomous pygidia of various sizes and from a variety of orders. Clearly, with so much variation in size and shape, no single consistent function can be assigned to the pygidium.