| 水螅纲(Hydrozoa)是刺胞动物中唯一包含有极少数淡水种类的一个纲,多数种在沿海生活。大多是附着在岩石、海草、贝壳上的小型水螅型动物,常被误认为是海藻。单体或群体生活,身体呈水螅型或少数种呈水母型,或水螅型与水母型同时存在于群体中,形成二态或多态现象,或是水螅型与水母型在生活周期中不同时期出现,形成世代交替。水螅纲的水螅型结构简单,没有口道,胃循环腔中也没有隔膜。水母型绝大多数具有缘膜,胃腔中没有刺细胞。水螅型及水母型的中胶层中均无细胞结构,生殖细胞均来源于表皮层(外胚层),个别种即使来源于内胚层,但最后仍在外胚层中发育成熟。约有3000种,可分为5目。 |
|
| |
 水螅纲的主要特征 水螅纲的主要特征 |
 多海产,少数淡水产 多海产,少数淡水产 |
| 大多有水螅型和水母型,有明显的世代交替,但以水螅型为主;少数种类仅有水螅型或水母型 |
| 单个或群体,有的种类有群体多态现象,即在同一群体中,有多种形态与机能的劳动分工的个体 |
| 水螅型结构较简单,只有简单的消化循环腔 |
| 水母型有缘膜,触手基部有平衡囊,内具平衡石,有平衡和感觉功能 |
| 刺细胞只存在于外胚层 |
| 生殖腺由外胚层产生 | |
| |
 花水母目 Anthomedusae 花水母目 Anthomedusae |
| 也称为裸芽目(Gymnoblastea),水螅型世代发达,体表围鞘不包围螅体,触手多呈棒状或丝状,常随年龄增长而数目增加。生殖腺附着在垂唇上,水母型存在或不存在,呈高杯状。如筒螅(Tubularia),其水母芽附着在亲本垂管上,受精卵也在亲本上发育,经浮浪幼虫及辐射幼虫后才离开亲本,附着后由单轴型出芽形成群体。多孔螅(Millepora)为水螅型群体,能分泌碳酸钙外骨骼,常与造礁珊瑚一起生活在热带海洋中。具营养体、指状体及水母芽体,这些个体死亡后在骨骼上留下无数小孔,故名多孔螅。伞形螅(Corymorpha)是呈单体水螅型,生殖芽分枝,附着在两圈触手之间,具自由的水母体阶段,水母体呈高杯状,仅有单个触手,行有性生殖。此外,真枝螅(Eudendrium)、贝螅(Hydractinia)、笔螅(Pennaria)及淡水水螅均属此目,它们的触手呈丝状。 |
| |
|

|
| 筒螅(Tubularia) | |
|
|

|
| 筒螅(Tubularia)水螅型群体 | |
|
|

|
| 外肋筒螅(Ectopleura crocea)水螅型群体 | | |
| |
|

|
| 多孔螅(Millepora)能分泌碳酸钙外骨骼 | |
|
|

|
| 多孔螅(Millepora)群体 | |
|
|

|
| 笔螅(Pennaria disticha) | | |
| |
|

|
| 真枝螅(Eudendrium) | |
|
|

|
| 伞形螅(Corymorpha nutans)水螅型个体 | |
|
|
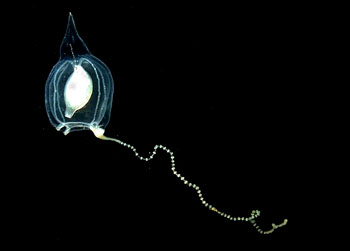
|
| 伞形螅(Corymorpha nutans)水母型个体 | | |
| |
|
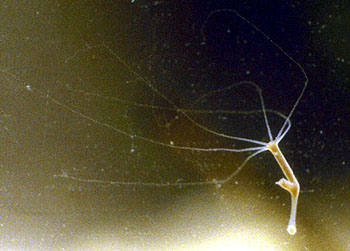
|
| 柄水螅(Pelmatohydra)属于淡水水螅,无水母型 | |
|
|
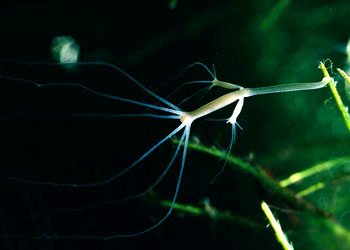
|
| 水螅(Hydra)属于淡水水螅,无水母型 | |
|
|

|
| 绿水螅(Chlorohydra)在内胚层内有绿藻共生,体呈绿色 | | |
| |
|

|
| 束状高手螅(Bougainvillia ramosa)水螅型群体 | |
|
|
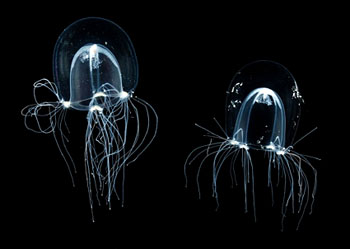
|
| 不列颠高手螅(Bougainvillia britannica) 水母型个体 | |
|
|

|
| 隔膜螅(Leuckartiara)水母型个体 | | |
| |
|

|
| 贝螅(Hydractinia)群体附着于寄居蟹寄居的贝壳外 | |
|
|

|
| 囊水螅(Euphysa)水母型个体 | |
|
|

|
| 全螅(Catablema nodulosa)水母型个体 | | |
| |
|

|
| 棍螅(Sarsia nipponica)水螅型 | |
|
|
|
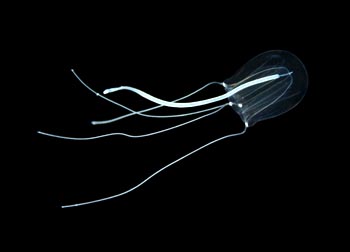
| 棍螅(Sarsia tubulosa)水母型个体 | |
|
|
|
| 芮氏螅(Rathkea)水母型个体 | | |
| |
| 管水母目 Siphonophora |
| 均为较大型的营漂浮生活的水母型群体。身体是由几种变态了的水螅型及水母型个体被共肉茎联结在一起,个体间紧密聚集,彼此分工组成一大型群体。一般根据浮囊体及游泳体的有无,可将管水母目分为3个亚目。钟泳亚目(Calycophora):没有浮囊体,仅有游泳体,例如五角水母(Muggiaea)。胞泳亚目(Physonectae):具浮囊体及游泳体,例如盛装水母(Agalma)。囊泳亚目(Cystonectae):有浮囊体、无游泳体,例如僧帽水母(Physalia),是成簇的水螅集合体,水螅体分3类,包括指状个体、生殖个体和营养个体,分管捕食、生殖和摄食。它的顶端气囊直径可达30cm,在气囊下仅留有茎的残基,有气腺可分泌气体进入气囊,靠气囊僧帽水母可以在海面随风逐流。它没有游泳体及叶状体,在大多数个体中发现有刺细胞,它所射出的蛋白质有很高毒性,螫人极痛,能引起严重反应,如发热、休克和心肺功能障碍。一般认为管水母类是来自硬水母,是由自由游泳的辐射幼虫阶段及成体水母型阶段合并,并逐渐分化而形成的多态群体。 |
| |
|

|
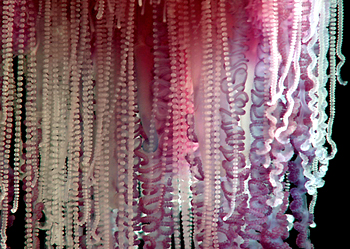
| 僧帽水母(Physalia)是成簇的水螅集合体 | |
|
|
|
| 僧帽水母(Physalia)刺细胞所射出的蛋白质有很高毒性 | |
|
|

|
| 僧帽水母(Physalia)顶端气囊可使其在海面随风飘流 | | |
| |
|

|
| 枝管水母 (Proboscidactyla) | |
|
|

|
| 异双生水母(Diphyes dispar) | |
|
|

|
| 五角水母(Muggiaea) | | |
| |
|

|
| 帆水母(Velella velella) | |
|
|

|
| 北极单板水母(Dimophyes arctica) | |
|
|

|
| 气囊水母(Physophora hydrostatica) | | |
| |
| 瘦水母目 Leptomedusae |
| 水螅型群体的围鞘包围螅体,营养体触手数目不随年龄而增加。大多不具自由生活的水母体。水母体多扁平盘形,多以假单轴型或复合型形成群体。如薮枝螅(Obelia)、海榧(Plumularia)、钟形螅(Campanularia)等。薮枝虫生活于浅海,固着在海藻、岩石或其他物体上,为一树枝状的水螅型群体。群体基部的构造很像植物的根,故称螅根(hydrorhiza),由螅体上生出很多直立的茎,称为螅茎(hydrocaulus)。螅茎上分出2种个体――水螅体(hydranth)与生殖体(gonangium)。整个群体外面,包围着由外胚层分泌的一层透明的角质膜,称围鞘(perisarc),具保护和支持的功能。水螅体主要管营养,其构造与水螅基本相同,有口及触手,触手是实心的,垂唇较水螅的长大,其外有一透明的杯形鞘,称为水螅鞘(hydrotheca)。生殖体无口及触手,只有一中空的轴,称为子茎(blas-tostylus),子茎的周围,有透明的瓶状鞘,称为生殖鞘(gonotheca)。生殖体能行无性生殖,其营养主要靠水螅体供给,因为水螅体和生殖体彼此由螅茎中的共肉(coenosarc)连接,整个群体的消化循环腔是相通连的。群体中任一水螅体捕食消化后,可通过消化循环腔输送给其它部分或其他个体。 生殖体成熟后,子茎以出芽的方法产生许多水母芽,水母芽成熟,脱离子茎,由生殖鞘顶端的开口出来,在海水中营自由生活,其结构较简单,很小,约1mm~2mm,体形如一圆伞,伞边缘生有很多细的触手(初生时16个),下伞面中央有一短的垂唇,口向内通到胃,再由胃伸出4个辐管,与伞边缘的环管相通。口、胃、辐管、环管构成水母的消化循环系。水螅水母特征之一是在伞下面边缘有一圈薄膜称为缘膜(velum),而薮枝虫的水母缘膜退化。在伞边缘有8个平衡囊,司平衡。在4条辐管上有四个由外胚层形成的精巢或卵巢(雌雄异体)。精卵成熟后在海水中受精。受精卵发育,以内移的方式形成实心的原肠胚,在其表面生有纤毛,能游动,称为浮浪幼虫(planula)。浮浪幼虫游动一段时期后,固着下来,以出芽的方式发育成水螅型的群体。 |
| |
|

|
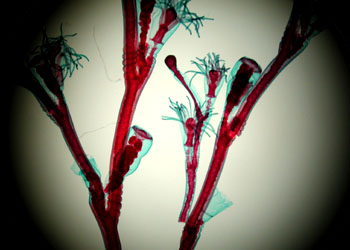
| 薮枝螅(Obelia)群体 | |
|
|
|
| 薮枝螅(Obelia)水螅型 | |
|
|
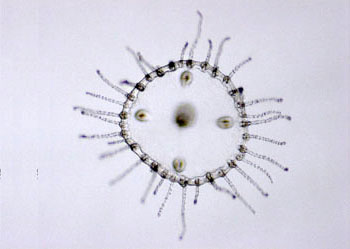
|
| 薮枝螅(Obelia)水母型 | | |
| |
|

|
| 海榧(Plumularia) | |
|
|

|
| 桧叶螅(Sertularella polyzonias) | |
|
|

|
| 桧叶螅(Sertularia argentea) | | |
| |
|

|
| 柏螅(Thuiaria thuja) | |
|
|

|
| 植丛管螅(Lafoea dumosa) | |
|
|

|
| 轮根茎螅(Rhizocaulus verticillatus) | | |
| |
|

|
| 瘤手水母(Tima formosa) | |
|
|

|
| 多管水母(Aequorea macrodactyla) | |
|
|

|
| 真瘤水母(Eutima gegenbauri) | | |
| |
| 淡水水母目 Limnomedusae |
| 生活史具有单体的小型的水螅型体和自由游泳的水母型体,但以水母型为主。水螅型体无围鞘,甚至无触手,单体常仅数毫米。水母体也为小型,都具缘膜,这是水螅水母的特征,具很多触手。例如桃花水母(Craspedacusta),是仅有的一种淡水生活的小型水母,生活在清洁的江河、湖泊之中,我国的嘉陵江上游有分布,为世界级濒危物种,是我国一级保护动物,有“水中大熊猫”之称。已记录11种,除索氏桃花水母(Craspedacusta sowerby)和日本的伊势桃花水母两种外,其余9种我国均产,如中华桃花水母(Craspedacusta sinensis)、楚雄桃花水母(Craspedacusta sowerbyi)等。水母体直径1.5~2cm,具有很多触手,缘膜很厚,其水螅型阶段仅有数毫米大小。钩手水母(Gonionemus)小型,广布于大西洋和印度洋-太平洋的浅海区,在暖海中密集成群。转钩手水母(Gonionemus vertens)刺细胞有毒,人被蛰后,皮肤灼痛,很快起疱,局部水肿,10~30分钟后感到乏力,开始麻木,四肢关节疼痛,呼吸困难并可能暂时停止,可致肝功能失常,急性症状持续4~5天。 |
| |
|

|
| 索氏桃花水母(Craspedacusta sowerby) | |
|
|
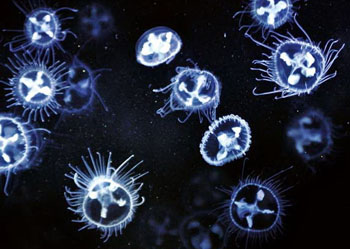
|
| 桃花水母(Craspedacusta)为我国一级保护动物 | |
|
|

|
| 转钩手水母(Gonionemus vertens)分布于浅海,有毒 | | |
| |
| 硬水母目 Trachylina |
| 生活史中没有水螅型,完全为水母型。体态属小型,生活在浅海到深海中。具或不具垂唇,生殖腺位于放射管下的表皮细胞间,发育中经浮浪幼虫及辐射幼虫再发育成水母型体。如壮丽水母(Aglaura)、三身翼水母(Geryonia)等。 |
| |
|

|
| 半口壮丽水母(Aglaura hemistoma) | |
|
|

|
| 三身翼水母(Geryonia) | |
|
|

|
| 银币水母(Porpita porpita) | | |
| |
| 水螅纲水螅型群体结构及其形成:水螅纲动物中除水螅,某些筒螅等极少数种为单体生活之外,其余绝大多数种类为群体生活。沿海常见的薮枝螅(Obelia)就是群体生活的代表,其群体呈树状,从几厘米到十几厘米,固着在岩石及海藻上。群体基部固着的部分呈水平方向生长,形成匍匐茎,也称螅根(hydrorhiza),由螅根上长出的直立茎称为螅茎(hydrocaulus),螅茎再分枝,分枝的末端长出螅体(hydranth)。薮枝螅的螅体有两种形态:种螅体有口有触手,具有取食与消化的机能称营养体(gastrozooid);另一种是无口无触手的棒形个体,个体的中央茎(也称子茎(blastost-yle))可以用出芽方式形成许多水母芽,这种个体称生殖体(gonozooid)。生殖体产生的水母芽离开母体后独立生活,即水母型体。水母体雌雄异体,行有性生殖,卵在水中受精后,形成浮浪幼虫,至此以后虫体营固着生活,以出芽方式形成新的群体。群体中营养体的口位于垂唇的顶端,不同的种类其触手的形态、数目及着生方式等均可不同。多数种类的触手是实心的,或呈棒状,端部具大量刺细胞聚集成球,原始种类均为棒状触手,且在营养体上散布,例如遍枝螅(Syncoryne)就是这种类型的触手。触手或为丝状的,刺细胞沿触手全长散布或规则排列。笔螅(Pennaria)具有这两种形态的触手;棒状触手在垂唇上散布,丝状触手在垂唇基部排成一圈。筒螅(Tubularia)全部为丝状触手,但排成两圈,分别位于垂唇的端部及基部。最进化的种类还是薮枝螅,触手全部为丝状,在垂唇基部排成一圈。群体的体壁也是由表皮层、胃层中间夹有中胶层所组成。个体之间以及基部相联的体壁称为共肉(coenosarc),胃循环腔在个体之间也是相互沟通的。营养体将消化后的营养物质经共肉及胃腔输送到整个群体。群体体壁的外表有一薄层表皮细胞分泌的支持物称为围鞘(perisarc),它是由几丁质及苯醌鞣化的蛋白质所组成,它对逐渐增大体积的群体起支持作用,对个体有保护的功能,所以可以看做是一种外骨骼。围鞘常在分枝处或个体基部形成一些环,起着加固作用。围鞘如果一直延伸到螅体周围,这种群体称为有围鞘的(thecate)群体,共围鞘随螅体而命名,如营养鞘(hydrotheca)及生殖鞘(gonotheca),薮枝螅便是有围鞘的群体。群体的围鞘如果仅包围螅根、螅茎及共肉部分,而不包围螅体,这种群体称无围鞘(athecate)的群体。例如笔螅、筒螅便是这种。水螅型群体的形成是以出芽方式进行,绝大多数种类芽体是由螅根或螅茎产生,很少种由营养体产生。群体形成的方式有以下4种:(1)螅根型(hydrorhizaltype),芽体单个的由螅根处产生,产生后垂直生长,每个芽体形成一个直立的螅体,这是一种原始的群体形成方式,多见于原始种类,例如贝螅(Hydractinia);(2)单轴型(monopodialtype),群体的生长带在第一个螅体的基部,所以第一个螅体的茎可以不断地生长延伸,螅茎延伸时,它的侧芽长出新的螅体,新螅体的基部也有生长带,它也不断地向前延伸,在延伸过程中再形成新的侧芽,如此重复形成群体,结果群体的主轴是由第一个螅体所形成,最老的螅体是在主轴的最顶端,例如真枝螅(Eud-endrium),这种群体形成方式在无围鞘的裸芽类(Gymnoblastic hydroids)中流行;(3)假单轴型(sympodialtype),第一个螅体的基部没有生长带,所以它不能向前延伸,而是通过出芽方式产生一个或多个侧芽,同样侧芽也不继续延伸,而是又产生新侧芽,新侧芽越过亲本芽体,因此群体的主轴是由许多螅体的茎联合组成),最年轻的个体在分枝的顶端,越老的个体越靠近群体的基部,这种生长类型在低等的有围鞘的被芽类(Calyptoblastic hydroids)中流行,例如Halecium;(4)复合型(compoundtype),这是在假单轴型的基础上又恢复到单轴型生长,螅茎及侧枝的末端不是螅体,而是生长点,因此它们可以不断地延伸,其芽枝及螅体均由侧面发生,每个侧枝来自一侧的生长点,这种生长方式似乎是最成功的。最大的水螅群体都是以这种方式形成,例如海榧(Plumularia)。这种类型出现在较高等的有围鞘的被芽类群体中。 |
| |
| 水螅纲水母型的退化:水螅纲中的一些种类具有自由游泳的水母体,薮枝螅就是这样,它的水螅型体以出芽方式形成独立的水母体,水母体又以有性生殖产生水螅型体,即具有世代交替现象。但多数种类并不产生自由游泳的水母体,其水母体世代是永远附着在亲本水螅群体上形成水母芽形式,也就是说它的水母型世代表现出不同程度的退化。例如贝螅的水母芽是永远附着在独立的螅茎上,遍枝螅及筒螅永远散布在营养体的垂唇上示群体的复合型形成方式。在真枝螅是发生在营养体的基部,总之这些水母芽表现出退化,它们没有口及胃腔、触手减少或消失、身体失去伞形。结果水母芽变成了囊状体,囊状体实际就是由外胚层形成一囊形的膜状结构,其中包含有大量的生殖细胞及内胚层形成的实心轴,水母形态已完全消失。在一种直杯螅(Orthopyrix),其生殖体也不形成水母芽,而形成一种端囊(acrocyst),其中也含有大量的生殖细胞,生殖细胞可以在端囊中受精并发育,直到形成浮浪幼虫之后,端囊才由生殖体中释放出去。尽管如此,附着的水母芽、囊状体、孢子囊都仍然代表了有性生殖的个体,只是它们的水母型形态表现出了退化而已。 |
| |
| 水螅纲的多态现象:水螅纲中许多营群体生活的种类都含有营养体与生殖体两种形态与机能完全不同的个体,这种现象称为二态现象。群体中如果包括两种以上不同形态与机能的个体,则称为多态现象。例如贝螅,群体中包含有四种不同形态的个体,即正常的具有触手,能捕食的营养体;没有口与触手,个体顶端具有大量刺细胞的指状体,个体或直立或弯曲,具有保护功能;还有刺状体(spinezooid),它的表皮内包有几丁质骨刺,具有支持及保护的功能;最后还有一种生殖体,它具有水母芽,但从不由群体中释放出能自由游泳的水母体,而是由水母芽产生精子或卵,并在海水中受精。而且有趣的是贝螅也具有很强的再生能力,再生时不同形态与机能的个体碎片只能再生出它原来类型的个体,这说明了群体中不同类型的个体在最初形成时,已经在发生学上被决定。所以贝螅的多态现象引起了发生学家极大的兴趣。多态现象在管水母类(Siphonophora)达到了最高发展程度,群体中的个体分化达到了七种之多,并且分属水螅型或水母型两种类型的个体。例如Agalma,群体中呈水螅型的个体有:(1)营养体,具触手,但触手细长,并缠绕有刺丝带,是群体中唯一可以取食、消化的一类个体;(2)指状体,没有口,但有大量的刺细胞分布在触手上,与营养体触手相似,这是一种保护性个体;(3)生殖体,无口、无触手、呈子囊状,可以通过出芽方式产生水母芽。群体中呈水母型的个体有(4)浮囊体(pneumatophore),这是一种变形的水母体,位于群体的顶端,中胶层不发达,在内、外胚层细胞之间形成一个大的气囊或气室,胃层细胞特化成腺细胞,由它产生二氧化碳或空气充满气室中,用以漂浮;(5)游泳体(nectophore),呈水母型,具缘膜,放射管及环管等结构,肌纤维也发达,但无口、无触手及垂唇,它的功能是使群体运动;(6)叶状体(phyllozooid),呈叶状或盔状,体型很小,与水母型相似,有很厚的胶质,在群体中担任保护与漂浮的机能;(7)水母型生殖体,个体呈退化的水母型结构,无口、无触手及感官,具有大量的生殖细胞,雌雄异体,但群体可以是雌雄同体,生殖细胞在释放之后个体死去。刺胞动物中多态现象的生物学意义可以认为是群体中个体之间的劳动分工,因为腔肠动物尚未出现器官系统来担任不同的生理机能,而是通过群体中个体的形态分化来担任不同的生理机能,这种分化是一种很原始的形式,这与腔肠动物的生活史及发展水平是密切相关的。在单态的种类,例如水螅、海葵等,身体呈水螅型,生活史很简单,即由水螅型体产生精、卵,并形成受精卵→浮浪幼虫→水螅型单体。生活史中没有水母型,有的种甚至没有浮浪幼虫期。在二态及多态生活的种类,生活史中有水螅型及水母型阶段,其水螅型行无性生殖,水母型或水母芽行有性生殖,其受精卵仍发育成水螅型体,其生活史成为水螅型体→水母型体或水母芽→受精卵→浮浪幼虫→水螅型体。结果形成无性世代与有性世代的交替现象。世代交替现象究竟是二态或多态现象的产物?还是由于腔肠动物生活史的改变造成了多态?按照前一种观点,原始的腔肠动物应是水螅型,它通过有性生殖,次生性的产生水母型体,因此形成了二态或多态。按照后一种观点认为,祖先腔肠动物是柔软的水母型,而水螅型是幼虫期的持续。这后一种观点在水螅纲硬水母目中得到了证实。因为硬水母类的受精卵在发育中通过浮浪幼虫及辐射幼虫(actinula)直接发育成水母型,其生活史是水母型体→受精卵→浮浪幼虫→辐射幼虫→水母体。一种小型的水螅Microhydra的个体发育正是如此,这说明辐射幼虫首先发展了固着习性,随着延伸固着期,停止发育成水母型,而最终发育成水螅型群体,所以生活史的改变形成了多态。这种现象与管水母类的发育也相符合,它的浮浪幼虫也是首先形成水母型体,即浮囊体、游泳体。因此世代交替的观点应该让位于水母体是成熟的刺胞动物,而水螅体是幼虫期的持续阶段的观点。当然这个论点是建筑在硬水母目是最原始的刺胞动物的基础之上的。 |
| |